
서 론
인류의 활동은 지구의 평균 표면온도의 상승을 가져왔고, 전세계 기후 변화의 주원인이 되고 있다. 이에 산업화 이전의 지구 평균 표면온도보다 1.5°C 이상 높아지지 않도록 국제 협약을 맺고 전세계적인 노력을 기울이고 있으나, 2018년 기준으로 이미 1°C까지 높아져 있는 것으로 보고되었다[1]. 최근 기후 관찰 결과에 따르면, 2023년 7월부터 2024년 6월까지 12개월 연속으로 1.5°C 상승했다고 한다[2]. 심각한 기후 위기에 직면한 지구 공동체는 다각도의 국제협력방안, 지속가능한 생산방법 개발 및 기후정책 변화 등을 모색하고 있는 중이다.
지구온난화의 원인인 온실가스에는 이산화탄소, 메탄, 아산화질소, 할로겐 가스 등이 있다. 이들 온실가스가 지구내 태양열을 흡수하고 우주로 빠져나가는 것을 늦추어, 지구 대기를 따뜻하게 유지시킨다. 2022년 전세계에서 53.8 Gt CO2eq의 온실가스가 배출되었으며, 이 중 이산화탄소가 71.6%를 차지하고 있고, 메탄이 21%, 아산화질소가 4.8%, 할로겐 가스가 2.6%로 구성되어 있다[3]. 전세계 온실가스에서 농업 배출원이 차지하는 비율은 12.3%이며, 그 중에서 가축과 분뇨에서 기인한 온실가스 발생은 절반 정도로 전체의 6.2%를 차지하고 있다[4]. 가축사육을 통해 발생하는 온실가스는 장내발효에 따라 발생하는 메탄과 분뇨처리에서 발생하는 메탄 및 아산화질소가 대부분이다[4]. 장내발효에 의해서 발생하는 메탄의 대부분은 반추동물에서 기인한다[5]. 국제연합은 전세계 인구가 2050년에 98억, 2100년에 112억에 이르고, 소고기 및 유제품의 소비량이 인구 증가에 따라 크게 높아질 것으로 예측했다[6]. 이에 따라 반추동물 사육의 증가로 인해 메탄 발생량이 늘어나고 지구 온난화를 더욱 가속화시킬 수 있다.
메탄은 지구온난화 지수가 이산화탄소보다 약 28배 더 높은 반면, 대기 중의 존재기간(lifetime)은 12년 정도로 이산화탄소의 300–1,000년보다 매우 짧다[7]. 이에 메탄 발생량을 저감하게 되면 지구온난화 완화에 즉각적인 효과가 가능하다. 반추동물의 장내발효 메탄 발생 저감방안에 대한 영양학적, 유전학적, 미생물학적 연구들이 지속적으로 진행되고 있다. 반추동물의 사료 조성을 변경하거나, 원료, 사료 첨가제를 적용하여 장내발효 메탄 발생을 감소시키는 방법은 비교적 용이하게 적용할 수 있고 빠른 적용 효과를 기대할 수 있다. 이에 사료조성 변경 및 원료, 사료 첨가제 적용에 대해 많은 연구가 진행되었고 상업화도 이뤄졌다[8,9]. 본 논문에서는 반추위 메탄 저감 효과가 상시적으로 10% 이상으로 반복 보고된 사료조성 변경 및 원료, 사료첨가제에 한해서 영양학적 저감 작용기전 및 연구 결과를 종합하여 살펴보고자 한다.
전세계 및 국내 온실가스와 메탄 발생 현황
유럽연합 집행위원회에서 최근 발표한 보고서에 따르면, 전세계 온실가스는 산업혁명 때부터 2019년까지 전세계 산업 및 경제의 발전에 따라 지속적으로 증가해왔다[3]. 하지만 COVID-19 팬데믹으로 인해 2020년에 온실가스 배출은 2019년 대비 3.7%가 감소하는 결과를 가져왔다. 그렇지만 팬데믹의 정점을 지나자 다시 온실가스 배출이 증가하여 2022년에 53.8 Gt CO2eq의 온실가스가 배출되었다. 이는 2019년 대비 2.3%, 2021년 대비 1.4%가 증가한 결과이다. 2022년에 배출된 53.8 Gt CO2eq의 온실가스 중 이산화탄소의 비율은 71.6%를 차지하고 있고, 메탄 21%, 아산화질소 4.8%, 할로겐 가스 2.6%로 구성되어 있다[3]. 또한 온실가스 배출원은 에너지 분야 58.2%, 운송 분야 13.4%, 농업 ․ 산림 ․ 토지 분야 18.3%, 산업공정 분야 6.6% 및 폐기물 분야 3.5%로 집계되었다[4]. 전체 온실가스 배출원에서 농업은 12.3%를 차지하는데, 그 중에서 가축과 분뇨에서 기인하는 직접적 가축사육 온실가스 발생은 절반 정도로 전체의 6.2%를 구성하고 있다.
대한민국 환경부 온실가스종합정보센터의 2021년 보고서에 따르면, 한국은 2021년에 676.6 Mt Co2eq의 온실가스를 배출하였으며, 이산화탄소가 91.3%로 대부분을 차지하고 있고 메탄이 4%, 할로겐 가스가 2.5%, 아산화질소가 2.2%로 구성되어 있다[10]. 한국의 온실가스 배출은 전세계 대비 에너지 분야와 운송 분야 합의 비율이 더 크고, 그 비율은 86.9%를 차지하고 있다. 이어서 산업공정 7.6%, 농업 3.2%, 폐기물 2.4% 순으로 구성되어 있다. 3.2%를 차지하는 농업 배출원은 전세계 농업 비율인 12.3%에 비해 크게 낮은 수준이다. 농업의 3.2%는 21.4 Mt Co2eq의 배출량을 의미하고, 그 중에서 가축사육에서 직접적으로 기인하는 온실가스 발생량은 9.7 Mt Co2eq로 농업의 45%를 구성하고 있으며, 한국의 온실가스 총배출량의 1.5%를 차지한다[10].
전세계 온실가스 중 이산화탄소 다음으로 두번째로 많은 비율로 존재하는 메탄은 3분의 2가 인류의 활동으로부터 발생하며, 메탄 발생량의 41%가 가축사육, 분뇨처리 및 쌀 생산을 포함하는 농업활동에서 기인한다[11]. 농업활동 중에서 가축사육에서 기인하는 메탄 발생은 73%이며, 대부분이 반추동물로서 그 비중은 비육우 35%, 젖소 30%, 면양 ․ 염소 ․ 버팔로에서 15% 정도이며, 나머지 20%는 비반추동물이 차지한다[5]. 국제연합의 최근 연구에 따르면 전세계 메탄 발생량은 2000년에서 2017년까지 9%가 증가했다[12]. 전세계 경제가 발전함에 따라 이 경향성도 계속 증가할 것으로 예상된다. 하지만 가축의 생산 효율 개선에 따른 메탄 발생량 감소는 유의미하게 크지 않아, 동물성 단백질 수요 확대로 증가한 메탄 발생량을 상쇄시키지 못하는 것으로 보고되었다[13].
2022 국가 온실가스 인벤토리 보고서에서 2020년 한국의 메탄 발생량은 27.4 Mt CO2eq이었다[14]. Table 1과 같이, 농업 분야가 43.2%를 차지하고, 폐기물 분야에서 32.1%, 에너지 분야에서 21.7%의 메탄을 배출했다. 가축의 장내발효를 통해 배출된 메탄은 4.7 Mt CO2eq로 전체의 17.3%를 구성하고 있으며, 농업 분야의 40%를 차지한다. 농업 분야에서 배출되는 메탄은 2000년에서 2020년까지 큰 변화가 없었다[14]. 그 이유는 가축사육 및 분뇨처리에서 배출되는 메탄은 가축사육두수의 증가에 따라 배출이 증가되었지만, 벼 재배, 농경지토양 및 작물잔사 소각에서 기인하는 메탄의 배출이 감소되어 서로 상쇄했기 때문이다.
Data from Greenhouse Gas Inventory and Research Center [14].
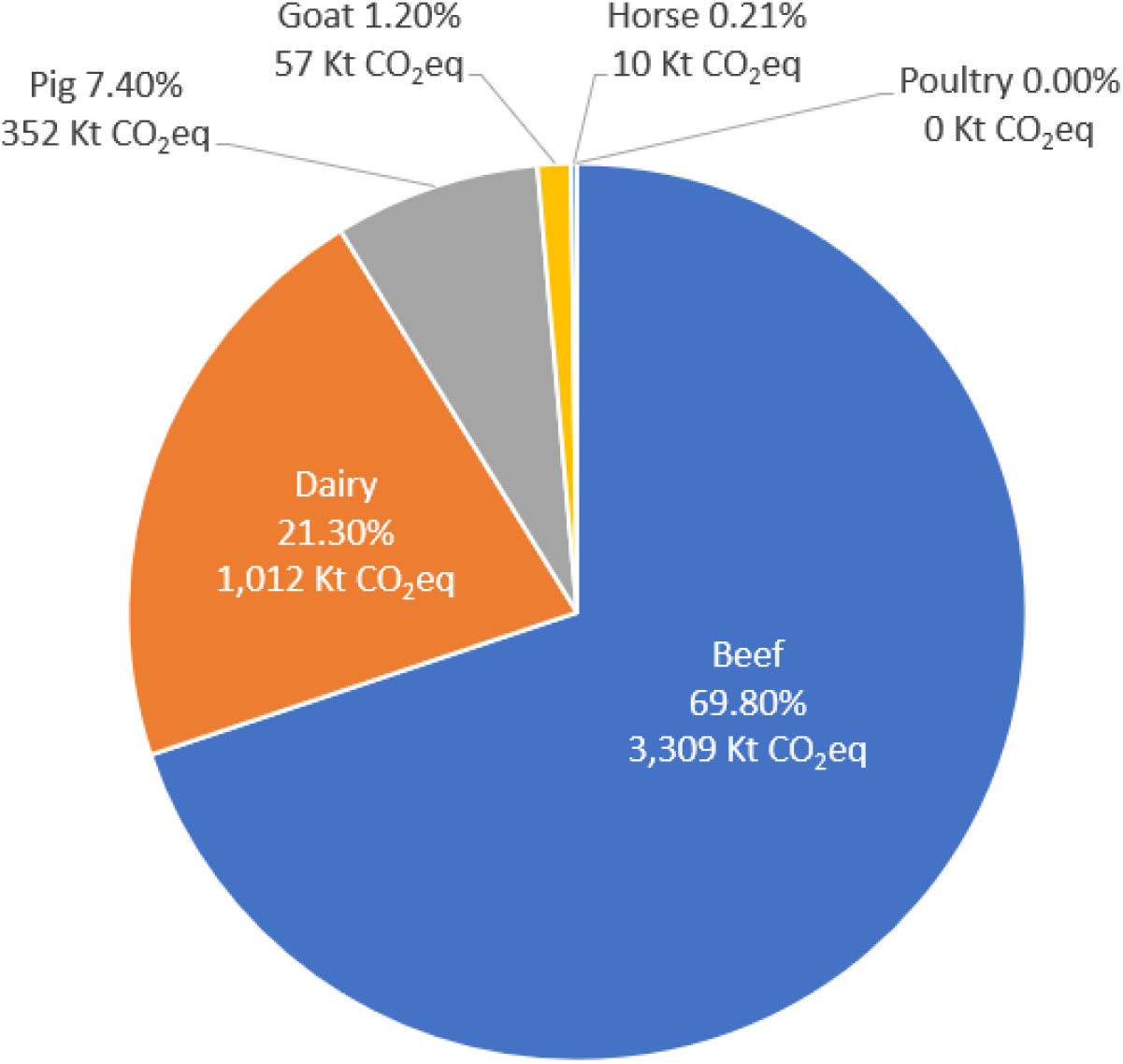
2020년 한국에서 가축의 장내발효를 통해 배출된 메탄은 4,743 Kt CO2eq이었다[14]. 이 중 반추동물이 92.3%를 차지하는데, 비육우가 69.8%, 젖소가 21.3%, 염소가 1.2%로 구성되어 있다(Fig. 1). 비반추동물 중에서는 돼지가 7.4%, 말이 0.21%를 차지한다. 이외에도 사슴과 면양이 메탄을 배출하나 미미한 부분을 차지하고 있다. 가금류는 장내발효를 통한 메탄을 배출하지 않는 것이 흥미롭다. 장내발효를 통해 배출된 메탄의 대부분이 반추동물에서 기인하기 때문에 반추동물의 사육두수 증가에 따라 메탄의 발생량은 또한 증가해왔다. 2000년에는 반추동물의 장내발효로부터 3,116 Mt CO2eq의 메탄이 배출된 반면, 2020년에는 4,381 Mt CO2eq의 메탄이 배출되어 42%가 증가했다[14]. 하지만 반추동물의 사육두수 정체에 따라 당분간은 국내 장내발효 메탄 배출량은 비슷한 수준을 유지할 것으로 보인다.
반추동물 장내발효에 의한 메탄 생성 과정
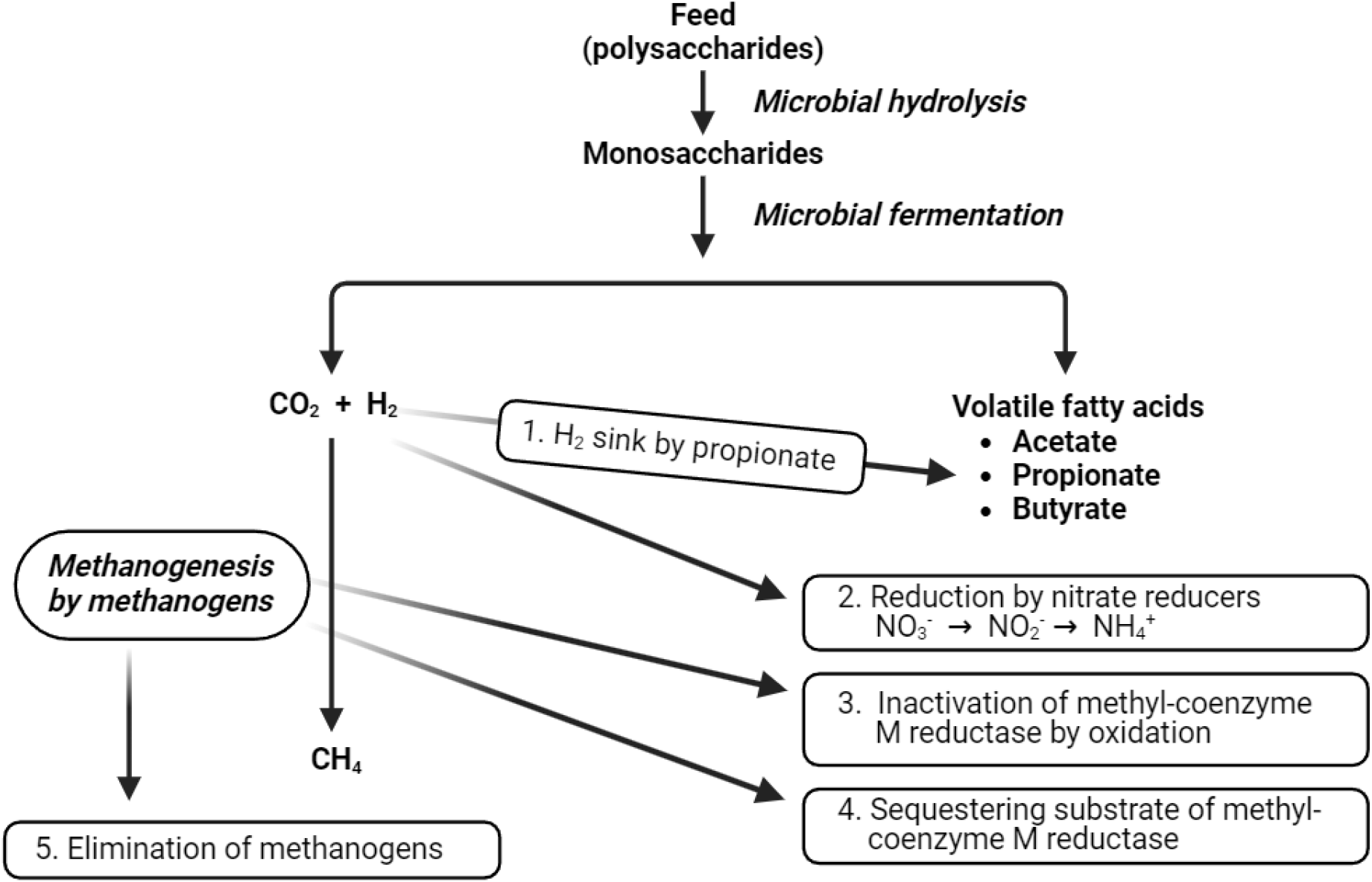
Fig. 2와 같이 반추위 미생물은 사료 내 다당류를 단당류로 가수분해하고 이어서 대사산물로 휘발성 지방산을 생성한다. 반추위 발효산물로 휘발성 지방산에 더해 이산화탄소, 수소분자, 발효열 및 미생물체 단백질 등을 생성한다. 반추위 메탄생성균이 이산화탄소와 수소분자를 결합시켜 메탄을 생성하거나, 메탄 파생물 또는 아세트산을 기질로 해서 메탄을 생성하기도 한다[15]. 다만 이산화탄소와 수소분자를 결합시켜 메탄을 생성하는 과정이 82%를 차지하여 반추동물의 주요 메탄 생산경로가 된다[16]. 이 과정을 통해 반추위 내 수소분자가 제거되어 반추위 발효가 원활히 진행될 수가 있다[17].
반추위 메탄생성균은 Wood–Ljungdahl pathway라 불리는 환원적 acetyl-CoA 경로를 통해 이산화탄소를 메탄으로 환원시키는데, 이는 혐기적 환경에서 탄소를 고정시켜 에너지를 재생시키는 중요한 과정이다[18]. 이 과정에서 전자 공여자로 주로 수소분자가 사용되기 때문에[19], 이산화탄소와 수소분자의 결합이 메탄 생성의 핵심이 된다. 반추위 메탄 생성 과정에서 methyl-coenzyme M reductase(MCR)라는 환원효소가 존재하는데, Wood–Ljungdahl pathway의 마지막 단계에서 MCR이 methyl thioether methyl-coenzyme M의 메칠기를 환원시켜 메탄으로 전환시킨다[20]. 이때 thiol coenzyme B를 전자 공여자로 사용한다. MCR은 반추위 메탄 생성 과정에서 유일하게 필요로 하는 효소이기 때문에, 이 효소를 불활성화 하거나 효소의 기질을 제거함으로써 메탄의 생성을 저해하는 시도가 연구되어 왔다[21,22].
반추위 생성 메탄의 영양학적 저감 작용기전
반추동물 장내발효에 의해 생성된 메탄을 감소시키는 것은 지구온난화 완화 전략에서 중요한 역할을 하기에 많은 연구자들이 영양학적, 유전학적, 미생물학적 분야에서 다양한 저감 전략을 연구해 왔다. 반추동물 사료조성의 변경 및 원료, 사료 첨가제의 적용은 비교적 용이한 적용 방법과 빠른 적용 효과를 가지고 있기에, 이러한 영양학적 저감 방법에 대해 연구 및 상업화가 많이 진행되었다. 하지만 메탄 저감 원료 및 사료 첨가제의 효능과 적용 가능성에 대해 체계적으로 비교 정리한 문헌이 없어서 소비자들의 혼란이 있었다. 이에 2021년 Global Research Alliance에서 11개 그룹의 원료 및 사료 첨가제의 메탄 저감 효능, 적용 가능성, 제약사항 등을 평가하여 제시한 바 있다[9]. 이 문헌에 따르면 3-nitrooxypropanol(3-NOP)과 홍조류(Asparagopsis)가 상시적으로 20% 이상의 반추위 메탄 저감이 가능하고, 질산염, 불포화지방산, 에센셜 오일은 10%–15% 정도의 저감이 가능한 것으로 보고되었다. 비섬유성 탄수화물의 비율을 높이는 사료조성의 변경 역시 10% 이상의 반추위 메탄을 감소시킨다[8]. 이에 본 논문에서는 반추위 메탄 저감 효과가 상시적으로 10% 이상으로 반복 보고된 사료조성 변경 및 원료, 사료 첨가제에 한해서 영양학적 저감 작용기전을 살펴보고자 한다.
Fig. 2와 같이 반추위 발효를 통해 발생한 수소분자가 이산화탄소와 결합하여 메탄을 형성하기 때문에, 수소분자의 대사경로를 전환시켜 이산화탄소와 결합하지 못하게 하면 메탄 발생이 감소하게 된다. 반추위 내 수소분자의 대사경로를 전환시키는 물질로는 프로피온산과 질산염이 있다. Fig. 2의 ①의 경로와 같이 반추위 내 휘발성 지방산 중 하나인 프로피온산은 수소 흡수원으로 작용한다. 이 때문에 프로피온산의 생성을 증가시키는 것은 메탄 발생을 저감시킬 수 있다[23]. 그리고 Fig. 2의 ②의 경로에서, 질산염은 반추위 내 수소분자와 결합하여 암모니아를 형성함으로써 수소분자의 대사경로를 전환시켜 메탄 발생을 감소시킨다[24].
반추위 메탄생성균에 의한 메탄 생성 과정의 마지막 단계는 MCR이라는 환원효소가 필수적으로 작용한다. 메탄생성균에 존재하는 MCR은 methyl thioether methyl-coenzyme M과 thiol coenzyme B을 기질로 하여 메탄으로 환원시키는 역할을 한다[20]. 이에 MCR을 불활성화 시키거나 MCR이 기질을 이용하지 못하게 함으로써 메탄생성 대사경로를 차단할 수 있다[21,22]. MCR을 불활성화 시키는 물질은 3-NOP라는 유기 화합물이 대표적이며, Fig. 2의 ③의 경로에서 메탄생성 대사경로를 차단한다[21]. MCR의 기질 이용을 방해하는 물질은 홍조류에 존재하는 bromoform이며, Fig. 2의 ④의 경로에서 메탄생성 대사경로를 차단한다[22].
반추위 메탄생성균은 메탄 파생물 또는 아세트산을 기질로 해서 메탄을 생성하기도 하지만, Fig. 2와 같이 주로 이산화탄소와 수소분자를 결합시켜 메탄을 생성한다. 이에 이산화탄소와 수소분자의 결합이 반추동물의 주요 메탄 생산경로가 된다[15]. Fig. 2의 ⑤의 경로에서 메탄생성균을 제거하면 반추위 메탄 생성을 감소시킬 수 있다. 대표적인 물질로 에센셜 오일과 불포화지방산이 이 경로에서 메탄을 저감시킨다. 에센셜 오일은 반추위 메탄생성균의 성장을 저해시켜 메탄 생성을 감소시킨다[25]. 불포화지방산은 메탄생성균에 독성으로 작용하거나 메탄생성균의 세포막을 파괴시켜 메탄 발생을 감소시킨다[26].
반추위 생성 메탄의 영양학적 저감 전략
반추위 발효에 의해 메탄이 발생되기 때문에, 일반적으로는 반추위의 소화 및 발효생리를 이용하여 발효산물을 조정함으로써 메탄 발생을 저감시킬 수 있다. 사료의 조성을 변경하여 발효산물을 조정하는 것은 적은 비용으로 반추위 메탄 생성을 감소시키는 방법으로 잘 알려져 있다[24]. 가장 일반적인 방법은 조사료의 종류 및 품질을 변경하거나 조농비율을 조정하는 것이다. Cellulose와 같은 구조탄수화물을 함유한 저질 조사료를 급여하였을 때는 메탄 발생량이 증가하고, 양질 조사료 및 농후사료와 같은 가용성 탄수화물이 많이 함유된 사료 급여 시에는 발생량이 감소하게 된다[27]. 가용성 탄수화물의 발효로 인해 더 많이 생성된 프로피온산이 수소 흡수원으로 작용하기 때문에, 메탄생성 경로에서 필요로 하는 수소분자를 감소시켜 메탄 발생을 저감시키는 것으로 알려져 있다[23].
반추위 메탄 발생에 영향을 주는 사료의 조성 변경 연구를 정리한 종설 논문[8]에 따르면, 사료의 조성 변경은 반추위 메탄을 0%에서 15%까지 저감시킬 수 있으며, 최대치인 15%는 에너지 보정 우유 kg당 2.25 g의 메탄을 줄일 수 있는 수준이다. 이 종설 논문에서 조사료의 품질을 높여서 가용성 탄수화물 발효를 더 유도하는 것은 최대 5%까지의 메탄 저감 효과를 보여줬다. 그리고 양질의 조사료로 대체하는 것도 비슷한 효과를 보였다. 또한, 곡물을 가공하여 급여할 때도 더 많은 프로피온산이 생성되어 1%–2.5%의 메탄 저감이 가능했다. 농후사료를 더 급여할 수 있도록 조농비율을 변경하는 것은 최대 15%까지 메탄 저감이 가능하였고, 비섬유성 탄수화물 1% 증가 시 메탄 2%가 감소하는 상관관계가 있음을 지적하였다[8]. 다만 농후사료의 급여 수준을 크게 증가시키는 것은 반추위 내 젖산 및 휘발성 지방산의 생성을 증가시켜 아급성 산독증과 같은 대사성 질병을 일으키기 때문에 주의가 요구된다.
질산염은 반추위 발효를 통해 발생한 수소분자가 이산화탄소와 결합하여 메탄을 형성하지 못하도록 수소분자의 대사경로를 전환시켜 메탄 생성량을 감소시키는 대표적인 물질이다. 일반적으로 질산염은 칼슘과 결합한 이온성 화합물 형태로 반추동물에 급여한다. 질산암모늄의 경우 폭발성 물질이기 때문에, 사료용 원료로는 주로 질산칼슘이 이용된다[9].
질산염은 수용성이며 반추위 혐기성 환경에서 빠르게 암모니아로 환원되는 성질을 가지고 있다. 이 환원 과정에서 반추위 수소분자가 이용되면서 수소 이용의 경쟁관계에 있는 메탄이 적게 생산된다[24]. 반추위 메탄 생산에 미치는 질산염 효과에 대한 연구는 2010년 이후 면양, 비육우, 젖소에 대해 광범위하게 이뤄졌다(Table 2). 4건의 면양 연구에서 사료 건물 2.5%의 질산염 급여는 대조구 대비 30%의 메탄을 감소시키는 것으로 보고되었다. 또한 8건의 비육우 연구에서는 사료 건물 2.2%의 질산염 급여로 21%의 메탄이 감소되었다. 7건의 젖소 연구에서도 이와 유사하게, 사료 건물 2.1%의 질산염을 급여했을 때 평균 22%의 메탄이 감소되었다.
| Study no. | Authors | Year | Animal | Nitrate level (% of DMI) | Methane reduction (% of control) | References |
|---|---|---|---|---|---|---|
| 1 | Van Zijderveld et al. | 2010 | Sheep | 2.6 | 32 | [28] |
| 2 | Nolan et al. | 2010 | Sheep | 2.5 | 23 | [29] |
| 3 | Li et al. | 2012 | Sheep | 2.3 | 34 | [30] |
| 4 | El-Zaiat et al. | 2014 | Sheep | 2.7 | 33 | [31] |
| 5 | Hulshof et al. | 2012 | Beef cattle | 2.2 | 32 | [32] |
| 6 | Leng et al. | 2012 | Beef cattle | 1.8 | 21 | [33] |
| 7 | Newbold et al. | 2012 | Beef cattle | 0.6–3.0 | 9–28 | [34] |
| 8 | Troy et al. | 2015 | Beef cattle | 2.2 | 12 | [35] |
| 9 | Lee et al. | 2015 | Beef cattle | 0.2–2.5 | 3–21 | [36] |
| 10 | Duthie et al. | 2018 | Beef cattle | 2.2 | 11 | [37] |
| 11 | Rebelo et al. | 2019 | Beef cattle | 1.4 | 19 | [38] |
| 12 | Villar et al. | 2020 | Beef cattle | 2 | 26 | [39] |
| 13 | Van Zijderveld et al. | 2011 | Dairy cow | 2.1 | 16 | [40] |
| 14 | Guyader et al. | 2014 | Dairy cow | 2.3 | 22 | [41] |
| 15 | Guyader et al. | 2015 | Dry cow | 2.3 | 28 | [42] |
| 16 | Veneman et al. | 2015 | Dairy cow | 2 | 22 | [43] |
| 17 | Olijhoek et al. | 2016 | Dairy cow | 0.5–2.1 | 6–23 | [44] |
| 18 | Klop et al. | 2016 | Dairy cow | 2.1 | 28 | [45] |
| 19 | Meller et al. | 2019 | Dairy cow | 1.5 | 17 | [46] |
질산염의 메탄 저감 연구를 정리한 종설논문[47]에 의하면, 질산염 1% 급여는 일반적으로 11.4%의 메탄을 감소시키고, 지효성(slow-release) 질산염 1% 급여는 13.2%의 메탄을 감소시킨다. 지효성 질산염 급여 시 분변에서 질산염이 발견되는 것으로 봤을 때, 지효성 질산염이 반추위 내에서 충분히 방출되지 않았고, 이로 인해 메탄 저감 효과가 기대보다 크지 않는 것으로 보인다[48].
질산염의 급여로 생성된 암모니아는 반추위 미생물체 단백질 합성의 질소원으로 사용될 수 있다. 이에 질산염을 적용하는 사료의 조성에서 요소와 같은 비단백태질소화합물의 수준을 감량 조절해야 하고, 이를 통해 사료비를 절감시킬 수 있다. 과량의 질산염을 급여하면 질산염 환원 과정에서 생성되는 중간 대사산물인 아질산염이 반추위 내에서 축적되고 혈액으로 진입하여 독성작용을 발현할 수도 있다[49]. 이에 질산염은 사료 건물의 2%를 넘지 않는 것이 추천된다[47].
반추위 내 메탄생성 대사경로에서 MCR을 불활성화시키는 물질에는 할로겐 화합물, 술폰화 화합물, 클로로폼, 니트로 유도체 등이 있다[50,51]. 니트로 유도체인 3-NOP는 methyl-coenzyme M과 유사한 화학적 구조를 가지고 있어서, methyl-coenzyme M의 자리를 차지하고 MCR에 결합을 하게 된다. 이 과정에서 메탄은 생성되지 않으며, 3-NOP가 MCR을 산화시켜 더 이상 methyl-coenzyme M과 결합을 할 수 없게 변형시키게 된다[21]. 이러한 기전에 따라 3-NOP는 반추위 내에서 메탄 생성을 억제할 수 있다.
니트로 유도체로부터 니트로 독소가 발생하지만 반추위 미생물은 내성을 가지는 것으로 보인다. 착유우와 비육우를 대상으로 한 in vivo 급여 실험[52,53]에서, 건물섭취량, 유량 및 소화율은 대조구와 동일했으며, 일당증체량은 오히려 높아진 것을 확인할 수 있기 때문이다. Table 3에서 정리한 바와 같이, 3-NOP 급여에 따른 반추위 메탄 생성은 사료조성, 급여 기간 및 방법에 따라 20%에서 60%까지 감소되는 결과를 가져왔다. 이들 연구에서 3-NOP의 평균 급여량은 123 mg/kg DMI였으며, 이는 평균적으로 비육우에서 23%, 착유우에서 39%의 메탄을 감소시켰다. 비육우와 착유우 간 메탄 저감의 차이는 사료 조성의 차이에서 기인한 것으로 보인다. 착유우 사료와 같이 조사료의 비율이 높은 사료 급여 시 반추위 발효에서 메탄이 더 생성되어[54], 3-NOP가 착유우에서 더 많은 메탄 생성 경로를 차단했기 때문이다. 우사에서 사육하는 비육우 및 착유우에 대한 연구가 많은 반면, 방목하는 반추동물에 대한 연구가 전무하기 때문에 방목에 대한 추가적인 연구가 요구된다.
| Study no. | Authors | Year | Animal | 3-NOP level (mg/kg DMI) | Methane reduction (% of control) | References |
|---|---|---|---|---|---|---|
| 1 | Martínez-Fernández et al. | 2014 | Sheep | 0, 111.2 | 16–19 | [55] |
| 2 | Romero-Perez et al. | 2014 | Beef cattle | 0, 47.4, 143.6, 304.9 | 4–38 | [56] |
| 3 | Romero-Perez et al. | 2015 | Beef cattle | 0, 280.1 | 59 | [53] |
| 4 | Vyas et al. | 2016 | Beef cattle | 0, 100, 200 | 17–38 | [57] |
| 5 | Vyas et al. | 2018 | Beef cattle | 0, 50, 75, 100, 150, 200 | 18-10 | [58] |
| 6 | Vyas et al. | 2018 | Beef cattle | 0, 125, 200 | 41–42 | [59] |
| 7 | Martinez-Fernandez et al. | 2018 | Beef cattle | 0, 337.8 | 30 | [51] |
| 8 | Kim et al. | 2019 | Beef cattle | 0, 100 | 17 | [60] |
| 9 | Haisan et al. | 2014 | Dairy cow | 0, 129.5 | 60 | [61] |
| 10 | Reynolds et al. | 2014 | Dairy cow | 0, 26.6, 135.1 | 10-7 | [62] |
| 11 | Hristov et al. | 2015 | Dairy cow | 0, 40.0, 60.0, 80.0 | 24–33 | [63] |
| 12 | Lopes et al. | 2016 | Dairy cow | 0, 60.0 | 31 | [52] |
| 13 | Haisan et al. | 2017 | Dairy cow | 0, 68.3, 132.3 | 23–37 | [64] |
| 14 | Van Wesemael et al. | 2019 | Dairy cow | 0, 71.7, 75.1 | 23–28 | [65] |
니트로 유도체의 적용 이후 시간이 지남에 따라 점차 반추위 내성 미생물이 불내성 미생물로 바뀌게 되면서 메탄 저감 효과는 감소하는 것으로 보고되었다[66]. 또한 니트로 유도체로 인해 상당한 양의 메탄 생성이 감소되면, 반추위 내 수소가 축적이 되고 이는 장기적으로 반추위 발효에 부정적인 영향을 가져올 수도 있다[17]. 하지만 많은 연구에서 시간이 지남에 따라 메탄 저감 효과의 감소가 관찰되지 않기도 하였다. 따라서 3-NOP의 메탄 저감 효과가 장기간 유지되는지 확인하기 위해, 몇 년에 걸친 장기 급여 실험 또는 여러 산차에 걸친 착유우 실험이 향후 진행되어야 할 것이다.
반추위 내 메탄생성 대사경로에서는 MCR의 기질을 차단하여 메탄이 생성되지 않도록 할 수 있다. 대표적인 물질이 홍조류에 존재하는 bromoform이다. Bromoform은 할로겐 화합물 메탄 유사체로서, MCR의 기질이 되는 보결분자단(prosthetic group)과 결합하여 MCR이 기질을 이용하지 못하게 막는 역할을 한다[22]. 이에 따라 홍조류를 반추동물에 급여하게 되면 메탄 생성이 감소하는 것으로 알려져 있다.
Table 4에서 정리한 바와 같이, 홍조류 급여에 따른 반추위 메탄 생성은 사료 조성, 급여 방법, 품종 및 사육조건에 따라 10%에서 98%까지 감소되는 결과를 가져왔다. 홍조류 급여량에 따른 메탄 생성 저감율이 일정치 않은데, 이는 홍조류에 존재하는 bromoform의 함량이 유전적, 환경적 조건에 따라 자연적으로 다양하게 분포하기 때문이다[67]. 홍조류의 급여량을 사료 건물섭취량의 1%를 넘게 설정할 경우 섭취량의 저하가 관찰되었기에, 건물섭취량의 1%를 넘지 않는 것을 권장하고 있다[9]. 홍조류의 급여가 면양, 비육우, 착유우의 메탄 생성 및 생산성에 미치는 영향을 정리한 종설논문[67]에 따르면, 홍조류 급여로 반추위 메탄 저감 효과는 인정되지만, 반추동물의 사료섭취량, 반추위 휘발성 지방산 및 유고형분의 감소 등의 부정적인 생산성이 수반될 수 있어 홍조류 급여에 주의가 요구된다.
| Study no. | Authors | Year | Animal | Seaweed inclusion rate (% of DMI) | Methane reduction (% of control) | References |
|---|---|---|---|---|---|---|
| 1 | Li et al. | 2016 | Sheep | 0.5 / 1 / 2 / 3 | 6 / 19 / 25 / 29 | [68] |
| 2 | Kinley et al. | 2020 | Beef cattle | 0.05 / 0.10 / 0.20 | 9 / 3 / 98 | [69] |
| 3 | Roque et al. | 2021 | Beef cattle | 0.5 | 74.9 | [70] |
| 4 | Roque et al. | 2019 | Dairy cow | 0.5 / 1 | 26.4 / 67.2 | [71] |
| 5 | Stefenoni et al. | 2021 | Dairy cow | 0.5 | 29 | [72] |
| 6 | Alvarez-Hess et al. | 2023 | Dairy cow | 0.56 / 0.64 | 44 / 39 | [73] |
생태계에 자연적으로 존재하는 홍조류를 수확하는 것은 많은 비용과 노동력을 필요로 한다. 홍조류 내 bromoform은 빠르게 대사되는 특징을 가지고 있어서, 시간이 흐름에 따라 분해되어 불활성화 된다[72]. 이에 수확 즉시 동결건조 하여 보관 후 사료첨가제로 제조해야 하는데, 처리비용이 추가로 높아지는 단점이 있다. 홍조류를 육상에서 배양하고자 하는 시도도 있으나, 복잡한 홍조류 번식 특성 때문에 배양 및 대량 생산이 어려운 것으로 보인다[67]. 홍조류 급여에 따른 반추동물의 잠재적인 건강 문제도 우려된다. 반추동물에 홍조류 급여 시 반추위 염증 및 궤양이 발견되었다고 보고된 바 있고[67], bromoform이 인간 및 가축에게 발암물질로 작용할 수 있다고도 알려져 있다[74]. 이에 반추동물의 건강에 부정적인 영향을 주지 않는 수준에서 홍조류 급여량을 설정해야 한다.
식물유래 천연자원 물질은 장내 발효 조절을 통한 반추동물 생산성 개선 목적으로 많은 연구가 되었다. 유럽연합이 2006년에 성장촉진제로서의 항생제 사용을 금지하면서, 반추위 발효 효율 개선 및 메탄 생성 저감 목적으로 식물유래 천연자원 물질의 연구가 활발해졌다[75]. 이들 연구에서 반추위 메탄 생성 저감 효과가 뚜렷이 관찰된 식물유래 천연자원 물질로는 에센셜 오일이 대표적이다. 에센셜 오일은 식물이 천연적으로 생산하는 2차 대사산물로서 지방산이 아닌 소수성 화합물 형태로 존재한다. 동물에 필수 영양소는 아니지만 식물의 필수성분을 추출했다는 의미에서 에센셜 오일로 불리고 있다.
많은 연구에서 에센셜 오일의 항균 효과가 인정되었으며 인간 및 동물이 안전하게 섭취할 수 있는 것으로 알려져 있다[76]. 에센셜 오일은 반추위 메탄생성균의 성장을 직접적으로 저해시켜 이들을 제거함으로써 메탄 생성을 감소시킨다[25]. 하지만 에센셜 오일 종류에 따라서 반추위 미생물들이 다양하게 반응을 하기 때문에, 모든 에센셜 오일이 메탄생성균의 성장을 방해하는 것은 아니다. 일반적으로 garlic, thyme, rosemary, oregano, clove, eucalyptus, lavender, peppermint에서 추출한 에센셜 오일이 반추위 메탄 생성을 감소시킨 것으로 알려져 있다[5]. 에센셜 오일의 반추위 메탄 생성 저감 효과에 대한 in vitro 실험 결과들을 정리한 종설 논문[5]에 의하면, garlic은 73%–91%, thyme은 30%, rosemary는 20%, oregano는 11%–87%, clove는 34%, eucalyptus는 85%, lavender는 54%–60%, peppermint는 30%–52%의 반추위 메탄이 대조구 대비 감소되었다.
그동안 에센셜 오일을 이용한 많은 연구들이 수행되었지만, 대부분이 in vitro에 의한 실험실적 방법이고 일부 in vivo 실험도 단기간의 실험에 국한되어 있다. 반추동물에 급여하여 장기간 지속적인 효과를 확인한 연구는 거의 없는 것으로 보인다. 에센셜 오일의 장기간 급여로 인해 메탄생성균이 저해가 되면 반추위 내에 수소분자가 축적이 된다. 축적된 수소분자가 반추위 내 pH를 저하시켜 발효를 저해할 수 있기 때문에[17], 장기간 급여로 반추위 발효가 저해되는지 확인이 필요하다. 그나마 in vitro 실험에서도 동일한 에센셜 오일이 일관되지 않은 연구결과를 보이기도 했다. 이는 에센셜 오일의 첨가량이 다른 점도 있지만, 식물의 품종, 유전력, 수확시기, 추출 방법 등에 따라 같은 에센셜 오일이라도 주요성분의 함량 또는 효능이 차이가 날 수도 있기 때문이다[9]. 이러한 이유로 에센셜 오일을 이용하여 메탄 저감제로 상품화된 제품은 소수에 불과하다.
Beauchemin et al.의 연구에 따르면 사료 건물 kg당 34 g의 지방을 공급했을 때 평균 14%의 반추위 메탄을 감소시켰고, 장기간 급여 시에도 지속적으로 메탄을 감소시켰기 때문에 사료지방은 효과적인 메탄 저감제로 볼 수 있다[77]. 하지만 지방급여의 효력은 지방 형태, 급여량, 포화도, 지방산 쇄길이 등에 따라 달라지는 것을 확인할 수 있다[78]. 이에 반해 불포화지방산의 급여는 비교적 일관적으로 반추위 메탄 생성을 감소시킨다[78]. 이는 반추위 내 불포화지방산이 메탄생성균에 독성으로 작용하거나 메탄생성균의 세포막을 파괴함으로써, 메탄생성균의 감소와 함께 메탄 발생을 감소시키기 때문이다[26].
불포화지방산은 종실 또는 식물성 정제지방에 다량 존재하며, 인간 또는 동물의 섭취에 해롭지 않기 때문에 적극적으로 고려되는 메탄 저감 전략이다. 전체 지방산의 85%가 불포화지방산인 crushed linseed의 급여는 in vivo 실험에서 반추위 메탄을 12% 감소시켰다[79]. 같은 실험에서 extruded linseed는 동량 급여 시 반추위 메탄을 38% 감소시켰다. Linseed의 항영양인자인 시안화물을 제거하기 위해 보통 extrusion 가공처리를 하는데, 이 과정에서 진행된 열처리로 인해 extruded linseed의 반추위 메탄생성균에 미치는 불포화지방산의 영향력이 확대되었기 때문이다[79]. 국내에서 홀스타인 거세우를 대상으로 한 호흡챔버 실험에서도 일일 1 kg의 extruded linseed 급여가 15%의 반추위 메탄을 감소시킨 것을 확인할 수 있다[80]. 한편 불포화지방의 과량 급여는 섬유소 소화율 및 건물섭취량을 감소시키고, 유지방 합성을 저해하며, 유량 및 증체량에 부정적인 영향을 줄 수 있다[81]. 따라서 동물의 생리적 성장 단계, 기초사료의 지방 조성 등에 따라 세심하게 불포화지방의 적용량을 결정해야 한다.
결 론
반추동물 사육은 전세계 및 국내 메탄 발생에 있어 가장 큰 배출원이다. 전세계 인구 증가에 따른 소고기와 유제품의 수요의 증가로 전세계 반추위 메탄 배출의 증가 역시 예상된다. 한국에서의 반추위 메탄 배출은 2000년 이후 현재까지 크게 증가하였고, 향후 비슷한 수준을 유지할 것으로 보인다. 메탄의 지구온난화 지수가 이산화탄소보다 약 28배 높은 반면 존재기간은 짧기 때문에, 메탄 발생량의 저감은 지구온난화 완화에 즉각적인 효과를 기대할 수 있다. 따라서 반추동물 사육에서 메탄 발생을 저감하는 것이 가장 효과적인 지구온난화 대응 전략이 될 수 있다. 반추동물 사료의 조성을 변경하거나, 원료 또는 사료 첨가제를 적용하여 장내발효 메탄 발생을 감소시키는 방법은 비교적 용이한 적용 방법 및 빠른 적용 효과로 인해 연구 및 상업화가 많이 진행된 메탄 저감 전략이다.
이들 연구에서 반추위 메탄이 상시적으로 10% 이상 저감된다고 반복 보고한 사례만을 취합하여 작용기전 중심으로 정리를 해보았을 때, 크게 수소분자와 이산화탄소의 결합 방해, 메탄생성경로 차단, 메탄생성균 제거 범주로 분류할 수 있다. 반추위 가용성 탄수화물의 발효에서 발생하는 프로피온산과 사료 첨가제로 급여한 질산염이 반추위 수소분자의 흡수원으로 작용한다. 결과적으로 이산화탄소와 수소분자의 결합을 방해하여 메탄 생성이 감소하게 된다. 사료 첨가제 형태의 3-NOP와 사료원료로서 홍조류는 반추위 내 메탄생성 대사경로 상 중요한 효소에 영향을 미쳐 메탄 생성을 차단한다. 에센셜 오일은 반추위 메탄생성균의 성장을 방해하고 불포화지방산은 독성으로 작용하거나 세포막을 파괴하는 방식으로 반추위 메탄생성균을 저해하여 메탄 생성을 감소시킨다.
이와 같이 사료조성 변경 및 원료, 사료 첨가제 적용으로 반추위 메탄 저감 효과가 상시적으로 10% 이상이라고 반복 보고된 사례가 있으니, 이를 활용하여 전세계 및 국내 반추위 메탄의 배출을 줄이는 노력이 필요하겠다. 이러한 영양학적 전략을 적용함에 있어 세밀한 조정을 하지 않으면 대사성 질병, 생산성 감소, 가축 건강 위해 등이 발생할 수 있다. 또한, 일부 원료 및 사료 첨가제는 장기간 급여 실험을 통해 안전성 확인이 필요한 경우도 있다. 이에 메탄 저감 효율이 우수한 영양학적 전략을 대상으로 안전하고 장기적으로 지속적인 효과 발현을 위해 세부적인 추가 연구가 요구된다.